细胞质遗传xibaozhiyichran
由细胞质内的遗传物质所控制的遗传现象称细胞质遗传,又称核外遗传、非孟德尔式遗传或母体遗传。细胞质遗传的特点有:(1)正反交结果不一致,杂种只表现母本性状,这是最早在高等植物中发现的判断染色体外遗传的依据。例如,1909年德国植物学家科伦斯发现在紫茉莉的杂交试验中,黄绿色叶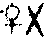 绿色叶的杂交子代都是黄绿色叶,而绿色叶♀×黄绿色叶的杂交子代都是绿色叶。这是由于真核生物的卵细胞除含细胞核外,还有大量的细胞质,而精子则除含细胞核外,仅有极少量的细胞质,所以合子的细胞质基本上来自卵细胞,当然合子的细胞质基因也基本上来自母本,那么,由这些细胞质基因控制的性状必然象母本,从而表现出母系遗传的特点。(2)细胞质遗传不遵循孟德尔的遗传规律,这是因为细胞分裂时,细胞质基因不是均等分配的,杂交后代一般不出现一定比例的分离现象 。(3)通过多次和雄亲本回交。
绿色叶的杂交子代都是黄绿色叶,而绿色叶♀×黄绿色叶的杂交子代都是绿色叶。这是由于真核生物的卵细胞除含细胞核外,还有大量的细胞质,而精子则除含细胞核外,仅有极少量的细胞质,所以合子的细胞质基本上来自卵细胞,当然合子的细胞质基因也基本上来自母本,那么,由这些细胞质基因控制的性状必然象母本,从而表现出母系遗传的特点。(2)细胞质遗传不遵循孟德尔的遗传规律,这是因为细胞分裂时,细胞质基因不是均等分配的,杂交后代一般不出现一定比例的分离现象 。(3)通过多次和雄亲本回交。
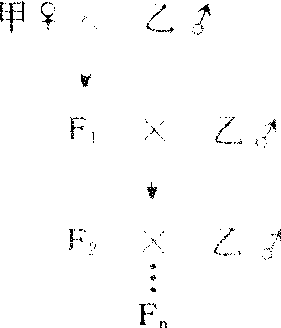
细胞质遗传cytoplasmic inheritance
染色体以外的遗传物质所决定的遗传现象。又称非孟德尔遗传或核外遗传。
1909年,重新发现“孟德尔定律”的法国植物学家科伦斯(C.E.Correns)报道紫茉莉 (Mirabilis ja-lapa)不符合孟德尔定律的遗传现象。花斑枝条(绿、白色镶嵌)♀×绿枝条♂的子一代有绿色、白色及花斑枝条;而绿枝条♀×花斑枝条♂的子一代全是绿枝条。类似的非孟德尔遗传现象在天竺葵 (Pelargonium Zo-nale)、月见草(Oenothera)、葫芦藓(Furnaria)及柳叶菜属(Epilobium)等植物中相继都有报道。研究者们提出细胞质基因组、质体基因及胞质基因等概念。1937年法国学者海蒂(P. H. Haitier)等报道果蝇对CO2敏感性的非孟德尔遗传。20世纪40年代相继在酿酒酵母、草履虫、大肠杆菌等微生物或原生生物中发现核外遗传现象。阐明这些遗传现象分别由细胞质中的线粒体、某些共生物及质粒控制。1963~1964年确证叶绿体及线粒体存在DNA,证实某些共生体及质粒亦具DNA性质,如果蝇的CO2敏感性因子σ和草履虫的卡巴粒,分别在细胞质中自成遗传体系。70年代绘制出质粒R100的遗传图。
正反杂交,是研究核遗传或细胞质遗传常用的方法。细胞质遗传时,正交及反交子一代总表现母本性状(如紫茉莉)、子二代表现不分离现象。子一代表现母本性状是因为双亲对子代提供相等的核物质,而细胞质只由母本提供。通过注射、接合转移(见细菌接合)等方法将某些生物(果蝇、大肠杆菌、草履虫等)的细胞质转移到遗传性不同的同类生物中,若受体获得供体的某些性状,该性状必属核外遗传。不断与父本回交,得到核置换材料(胞质来自母本,核来自父本)若仍表现母本某些性状,也是核外遗传的表现。显微操作进行核移植,从其性状表现亦可判断遗传方式。
生物有不少性状由细胞质基因或胞质基因与核基因共同作用决定。植物雄性不育特性在农作物杂种优势利用中的广泛应用就是例子(见雄性不育)。利用不同物种间叶绿体的摄取、转移可作为提高低光效作物(水稻、小麦)光合效率的手段。70年代报道烟草白花细胞原生质体摄取外源叶绿体后,能再生成正常烟草植株。矮牵牛及胡萝卜中亦有类似结果。
细胞质遗传
即“染色体外遗传”。
细胞质遗传cytoplasmic inheritance
又称染色体外遗传,核外遗传,非孟德尔式遗传或母体遗传。由细胞质基因决定性状表现的遗传现象。德国遗传学家柯伦斯(C.Correns)于1909年最先发现紫茉莉(Mirabilis jalapa)花斑植株的非孟德尔式遗传现象。现已证明该性状受质体基因控制。以后在粗糙面包霉、啤酒酵母、果蝇、草履虫以及许多植物中都发现这一现象。特点是:
❶正反交结果不同,F1均表现母本性状;
❷除花斑叶外,细胞质基因控制的性状一般不表现分离现象;
❸由附加体和共生体决定的性状,往往类似病毒的转导或感染;
❹细胞质基因可以通过连续多次回交,也可通过注射、细菌或草履虫的接合等方式转移;
❺细胞质基因对性状的作用有其自主遗传的特性,但某些性状受胞质基因和核基因的互作决定。应用重组分析、缺失分析、分子杂交和物理图谱分析等方法可以对细胞质基因进行基因定位。细胞质遗传已成为分子遗传学研究的一个重要项目。高等植物细胞质雄性不育基因的发现和研究,在杂种优势利用上有十分重要的价值。