质粒plasmid
染色体以外能自我复制的遗传因子。不具有胞外期,对寄主细胞来说是非必需的。质粒可能是DNA或RNA。质粒DNA不仅存在于细菌、蓝藻等原核生物中、在酵母、丝状真菌、植物、动物和人类等真核细胞中也有发现。除少数已鉴定出它们所编码的遗传性状以外,大多数是功能尚未清楚的隐蔽质粒。RNA质粒包括独立于寄主细胞染色体和细胞器基因组进行复制的非感染性RNA分子,以及具有蛋白质壳体的嗜杀双链RNA(ds RNA),具有外壳但不具感染能力的真菌病毒和植物隐蔽病毒的dsRNA。
理化性状 质粒DNA是共价闭合环状DNA分子(ccc DNA),其大小一般相当于染色体的0.1%~3%,通常分子量为2~150×106之间,而假单胞细菌等的质粒分子量可达300×106以上,足以编码上百个基因。不同种类的质粒在寄主中的拷贝数,或同一种质粒在不同寄主中的拷贝数都不相同,少则1~2个,多达几十个甚至上百个。质粒DNA嵌入溴化乙锭和吖啶类等呈平面结构的染料后,密度和构型改变,对酸碱及热的变性和复性作用,在流体力学上对切割的抵抗力不同于线状DNA和染色体DNA。最近在真菌和高等植物中发现称为附加体的线状双股DNA,能够整合进寄主染色体。RNA质粒通常是双链分子(ds RNA),存在于细胞质中的类病毒颗粒中,以及特化的脂质小囊中,少数RNA质粒也以单链形式(ssRNA)存在。
分类 一般按质粒的性质和特点进行归类。
根据质粒性质和转移特点,可分为接合型质粒和非接合型质粒,两者区别在于是否带有转移基因。接合型质粒具有在细胞间自我转移的能力,分子量大,每个细胞中质粒为1~3个拷贝。非接合型质粒分子量相对较小,在细胞中呈多拷贝。
按照质粒的复制特点,分为严紧型质粒和松弛型质粒。严紧型质粒复制伴随着染色体复制而进行,拷贝数少。松弛型质粒可在无寄主蛋白质合成的情况下复制。用氯霉素或其它物理因素处理某些松弛型复制的质粒细胞,可使拷贝数扩增到1000~3000个。
按表型效应,一般将大肠杆菌质粒分为致育因子(F因子),抗性因子和大肠杆菌素原因子。致育因子主要特征是接合作用,可通过性纤毛使原来不带致育因子的细菌带上该因子。抗性因子具有许多抗生素和某些重金属抗性特点,作为基因载体时是理想的标记选择。大肠杆菌素原因子编码大肠杆菌素,能特异性地杀死其它肠道细菌。携带该因子的菌株由于质粒本身编码一种对大肠杆菌素有免疫作用的蛋白质,而自身不受其伤害。
按照在共存的同一细菌中排斥同类质粒的能力,可分为相容性质粒和不相容性质粒。两种质粒被吸入同一细菌时,如果它们能一起复制并能共存,则它们是相容的,属于不同的不相容群。若两种质粒不能共存于同一细胞内,那么它们是不相容的,属于不相容群。
结构和功能 质粒一般分为必要区和非必要区。质粒含有某些染色体没有的基因,编码某些功能并非是细菌生存所必需的。
必要区 具有复制和调控系统,包括与质粒DNA复制、调控、不相容性等有关的基因。
非必要区 携带决定特殊表型的基因,目前已鉴定出质粒所控制的性状超过100种。如对抗菌素、重金属、阳离子、插入剂等抗性,分解芳香族化合物、产生抗菌素和细菌素等代谢能力,对其它生物的致病性和共生现象的控制。
与致病性的关系 质粒普遍存在于植物病原细菌和真菌中,不少病原菌的致病性与质粒及其编码基因控制的性状密切相关。
控制毒性 质粒不仅编码了病原菌产生激素、毒素等致病因子性状,也决定了致病因子尚不清楚的病原菌的致病功能。根癌土壤杆菌Ti质粒能诱发寄主细胞产生畸形的冠瘿瘤。Ti质粒中一段转移DNA(T-DNA)上致癌基因编码了生长素和细胞分裂素的合成,引起植物细胞激素失调,形成肿瘤。此外Ti质粒还决定肿瘤形态,寄主范围,冠瘿碱合成和利用,以及对农杆菌素K84敏感性的基因(见冠瘿瘤形成机制)。丁香假单胞菌致病变种的一些菌株产生丁香素、菜豆毒素、冠毒素以及烟草毒素,都是由质粒基因控制的。这些毒素引起植物叶片退绿和坏死。对玉米萎蔫欧文氏菌和青枯病假单胞菌进行质粒消除、缺失和治愈,突变体丧失或降低了致病性和毒性。许多病原真菌的致病性与双链RNA (ds RNA)质粒有关。维多利亚长蠕孢含有的沉降系数为145S类病毒颗粒中,具有4种ds RNA,与引起燕麦枯萎病有关。寄生隐丛赤壳(栗疫病菌)某些菌株含有一种控制减毒性状的ds RNA质粒,有毒菌株不含这类ds RNA。在立枯丝核菌中也发现质粒的存在与低毒力有关。
控制无毒性 在一些病原细菌中,控制对相应植物抗病品种无毒性的基因定位于质粒上。无毒基因可能涉及到病菌对特定植物的识别,对致病性和毒性起调节作用。辣椒斑点病黄单胞菌辣椒致病变种某些菌株具有一个45 kb大质粒,其中含有一个无毒基因,一个抗铜基因和三个拷贝的插入序列,该无毒基因与辣椒中相应的抗病基因互作,导致了不亲和反应。此外,丁香假单胞菌番茄致病变种和大豆致病变种的无毒基因也是质粒携带的。
生态适应和进化作用 质粒的存在使细菌对环境产生较强的适应能力。一是质粒在群体中的转移,使许多可资利用的基因得以在群体中传播和扩散,同时提高了细菌群体DNA复制的效果;二是质粒编码的性状,如产生细菌素和营养能力,增加了细菌在新生境中的适应和与其它细菌竞争的能力,对抗生素、重金属和紫外线辐射的抗性增加了细菌在不良环境中存活的机会。如玉米萎蔫欧文氏菌的质粒系统,有一套相互适应的基因,使其在周期性替换的小生境中保持稳定的表型。冠瘿土壤杆菌Ti质粒中T-DNA上具有的诱导植物合成冠瘿碱的基因,由于植物本身不能利用冠瘿碱,病菌驱使植物产生,供其作为唯一的氮碳源利用。
质粒基因的复杂性是细菌长期进化的产物。它们与染色体基因的相互作用,以及它们在群体中的传播,对加速宿主的进化具有重要作用。质粒不仅可作为转座因子的载体,引起宿主基因组的各种变化,而且还以转移和起动其它质粒基因和染色体基因,发生新的遗传交换、重组和变异。
应用 在分子植物病理学中,质粒可以在病原菌的致病性、其它有益性状以及分子操作中的载体方面加以研究应用。
致病性研究应用 鉴于病原菌的质粒与致病性的相关性,加强对质粒编码基因的类型,结构,表达和调控的研究,可以从分子水平阐明病菌的致病机制。
有益性状研究应用 已利用质粒编码的细菌素产生有益性状,应用在植物病害的生物防治中。如细菌素K84防治桃细菌性冠瘿病已获成功。
载体的应用 在分子操作中,质粒常作为基因的载体。但天然的质粒不一定是理想的载体,必须通过重组和改造来发展质粒载体。理想的质粒载体应该具有自我复制的复制子和高效表达的调控系统;具有多种限制性核酸内切酶的单一切点,切点最好位于易于检测的表型基因上;赋予宿主细胞易于检测的表型;分子量小,多拷贝;携带外源DNA幅度较宽。根癌土壤杆菌的Ti质粒经过去除T-DNA上的产生肿瘤基因后,作为基因载体广泛应用于植物的基因工程研究中。
质粒plasmid
染色体外的环状遗传因子。普遍存在于原核生物中,在一些真核生物中也有发现。1952年莱德伯格(J. Lederberg)首先提出质粒一词。某些既能独立地存在于细胞质中,又能整合在染色体上的质粒称为附加体。
细菌中的质粒根据其所含基因数量及功能分为大小两类。大质粒为100kb左右的双链DNA分子,足以编码上百个基因,但拷贝数很少。大质粒的复制与细胞分裂周期紧密相联,且其与自身复制有关的复制基因是成对地存在于整个基因组中,可通过细胞间的连接自主地控制其胞间的移动。控制转移的基因集中在基因组的一部分,大约占25kb,而另外70kb的DNA可带有70个基因,因而决定了它的多种性质。另一类为6~10kb的小质粒,只带6~10个基因,但拷贝数较多,每个细胞中有10~20个不带有转移的基因。但在可转移质粒存在时也能实现胞间的转移,质粒按其所携带基因的功能有抗药性质粒、致育性质粒、分解质粒等。一些以环状小分子形式存在而无明显表型效应的质粒被称为隐蔽性质粒。质粒的寄主性是与细菌的种类紧密相关的,但有些质粒则广泛存在于各类寄主中。
质粒不亲和群的分类是根据不同质粒在同一种寄主细胞中的共生能力而定。在相同寄主细胞中能够共同生存的质粒叫做质粒的亲和性,反之则为不亲和性。根据这项原则,在革兰阴性菌中发现有25种以上的质粒不亲和群。在革兰阳性菌中也有10多种 R质粒(抗性质粒)是最广泛存在于多种不同的不亲和群中的一类质粒。实验证明,质粒不亲和性决定于复制的控制。控制拷贝数的复制基因中一个碱基对的变化就可导致质粒由一种不亲和性转变为另一种。
在化疗及畜牧业中由于大量使用抗菌素,使带有多种抗性的细菌病原体大量增加。1967年,在日本痢疾流行时从患者肠道细菌中首次分离到了抗性质粒。细菌能抗各种抗菌素,如对青霉素、卡纳霉素、四环素等,而且也抗毒性重金属如汞、镉、银离子等。抗性质粒并不同时具有对以上抗菌素及重金属的抗性,但能同时抗4~6种抗菌素。这类抗性的生化机制主要有以下六种方式: ❶切断抗菌素中的特异化学键;
❷对抗菌素进行化学修饰导致失活;
❸合成新的对抗菌素不敏感的酶;
❹改变自身大分子上的作用基因;
❺降低细胞的通透性;
❻隔绝抗菌素,使其不能结合到作用位点。这样,同一种基因就能通过两种不同的机制来抵抗两种结构上完全无关的抗菌素。
在许多质粒中发现含有插入序列和转移遗传因子。插入序列存在于细菌基因组中,也可转移到其他染色体、质粒或病毒上去,其机理不同于正常的重组系统,它提供重组位点,能使各种基因融合在一起,使基因组带有更多的功能。因此插入序列对质粒与其他基因组进化中的遗传交流起到了媒介的作用。如大多数抗性基因位于两段方向相反的DNA重复序列中,这些序列与插入序列同源。转移遗传因子也像插入序列可插到新位点上。不同的是,它带有抗性基因外,还带有全部侧翼系列。它以极低的频率转移到其他基因组上去,使不同的质粒逐渐带有相同的抗性基因。对一些可转移和多抗性质粒的分子性质的研究表明,其形成是由掌管遗传转移的基因与抗性基因进入到含复制功能的小质粒后产生的。这样就保证了在染色体外遗传系统的进化中具有更优越的对寄主细胞的适应性。
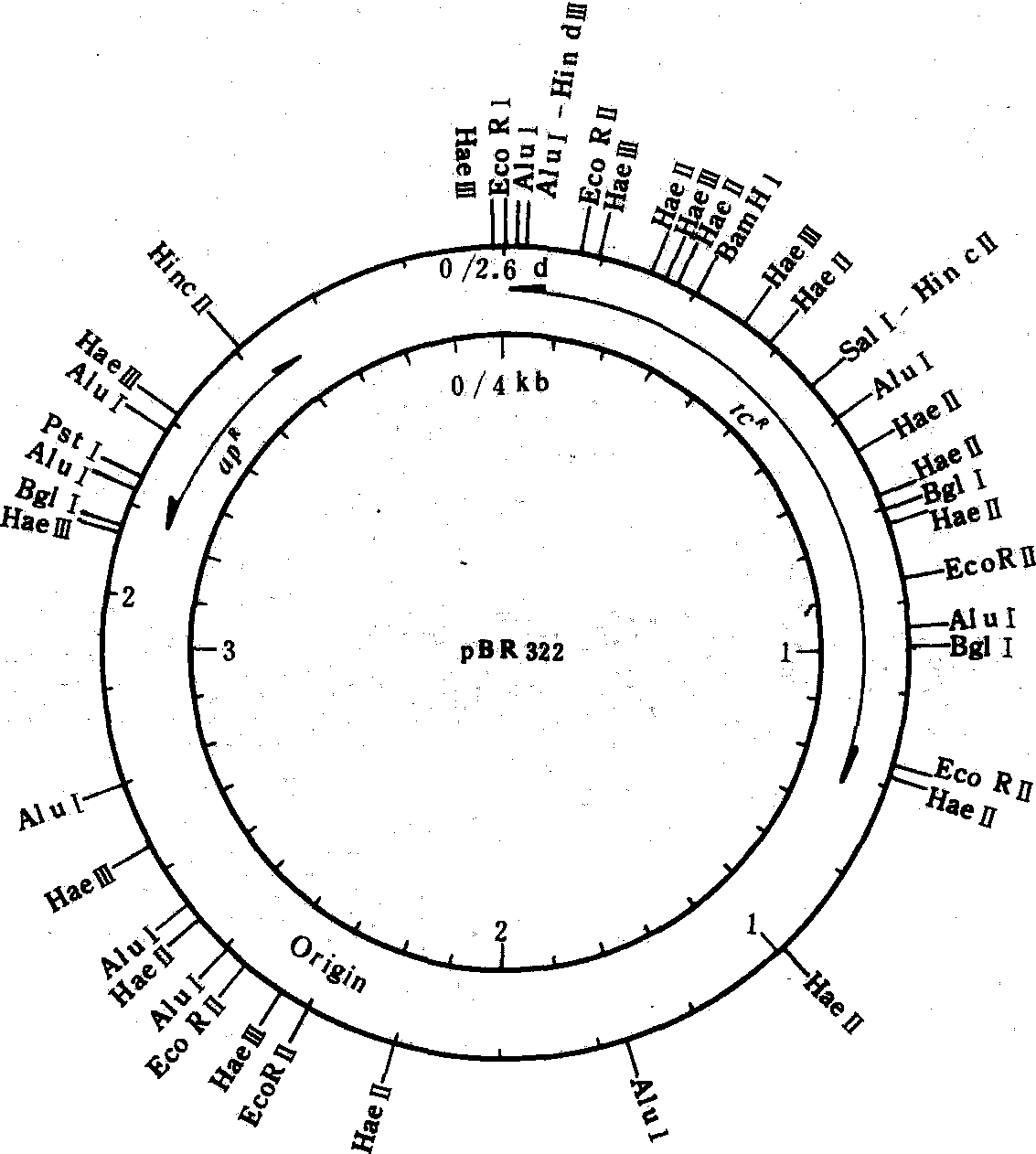
大肠杆菌质粒pBR322的结构示意图
里圈表示长度(kb),外圈示分子量(d)及各种限制酶的识别位点apR: 氨苄青霉素抗性基因; tcR: 四环素抗性基因; Origin: 复制起点
质粒在遗传工程中起着非常重要的作用。将一种生物的DNA片段与质粒连接起来,作为一种载体,通过转化引入细菌,这段DNA就会随着质粒的复制而复制,即随着细菌的分裂而扩增。这样就能得到这一DNA的大量纯制品,进行分子水平的研究。现在已制成了质粒及病毒载体,能在原核及真核细胞中高效地表达所克隆的基因。例如,PBR322是一种常用的大肠杆菌质粒(见图)。另外,还可以将两种宿主细胞的质粒连接成嵌合质粒作为穿梭质粒,能分别在两种宿主细胞中引入外源DNA并进行复制。
质粒
见“细胞”中的“质粒”。
质粒
染色体外的任何遗传结构。包括质体基因、附加体、潜伏性病毒和能复制的细胞器(叶绿体和线粒体)等。
质粒plasmid
存在于原核生物中的环形DNA结构。细菌细胞中染色体外能自主复制的遗传结构。例如F因子、R因子、colE1因子等等,可稳定遗传,但并非细菌生存所必不可少的成分。质粒DNA分子大小不等,一般仅及细菌染色体DNA分子的0.5%~3.0%,易从细菌内取出或引入。在细菌中的数目多少不一,严紧型质粒在细菌里只有1~2个,松弛型质粒可有20~60个之多。也有换入染色体与之同步复制的质粒,称附加体(episome)。一个质粒可有2~250基因,有特定的核酸内切酶切点,在不同基因间可以重组。可通过转化、转导和接合而转移,具备基因工程中作为基因载体的有利条件。实验室中应用最多的质粒载体有:pSC101 (抗四环素质粒),colE,(大肠杆菌素质粒)以及两者优点互补的共接质粒pSC134和其他众多经改造的优良质粒载体。高等植物基因工程中,Ti质粒是有希望的质粒载体。在理论上,质粒的研究涉及DNA复制、基因重组、生物进化等重要课题。由于抗病性质粒在致病菌中的传播,也使质粒成为流行病学研究的对象。(见“基因工程”)
质粒
质粒作为基本载体,在遗传工程中占有重要的位置。质粒是由环状双链的DNA分子组成。它大多存在于细菌的细胞质中,是细菌染色体以外的一种遗传系统。它能够独立复制自己,也可以和寄主细胞染色体协同进行。所以一个寄主细胞内可以含有许多质粒。质粒对细菌无伤害作用,但是当质粒进入细菌细胞质,细菌的表现性状就会有所改变。这是由于质粒既要从活细胞获得原料(核苷酸)来复制自己,又是靠活细胞的条件来表达自己的基因。
遗传工程中所用的质粒通常是独立的系统,不与细菌的染色体结合在一起。在采用质粒作为基因载体时,要分离出许多质粒,并通过加工,使他们便于和基因结合,容易进入寄主细胞,这样的质粒是一种具有新遗传特性的质粒。它一旦成功地进入活细胞,就会发挥其作用。