彩陶钵
新石器时代仰韶文化庙底沟类型。河南省陕县庙底沟村出土。庙底沟类型因首次在陕县庙底沟被发现而得名。典型器物有深腹、曲壁的碗和盆,双唇、小口尖底瓶等。此器宽口,腹部横鼓,愈到盆底愈小,形成高足。器表研磨光滑,以黑彩绘放射形的旋花纹。图案具有抽象性,笔触极为圆润流畅。这种动感极强的纹饰绘制在曲线突出的器物上,能增添韵律感,极引人瞩目。仰韶文化见“小口尖底红陶瓶”条目。
彩陶钵
新石器时代马家窑文化马家窑类型。高10.4厘米,口径29厘米。1975年甘肃省临夏水地陈家出土。甘肃省临夏回族自治州博物馆藏。这件器物内外均以黑彩装饰,器内纹饰更是生动。那飞舞的线条令人产生各种丰富的联想。例如,可以想象为对一个湖泊的概况描述。外层相互连接的弧线三角纹似荡漾着的水波拍击湖岸;内部的同心圆像风和日丽时的水面;中心的三组曲线象征不易觉察的水中暗流;点缀其间的黑点是湖中的各类生物。这件造型端庄规整的陶钵配以动感极强的纹饰,颇能够调动人们丰富的想象力,而令人难以忘怀。马家窑文化见“束腰彩陶罐”条目。
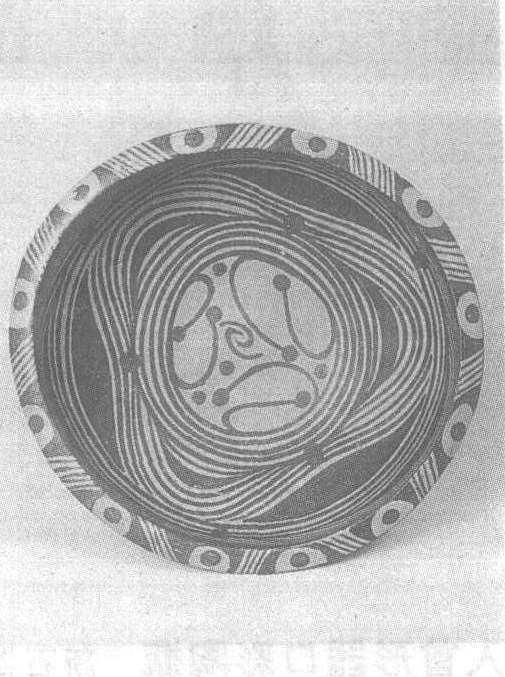
彩陶钵
战国。通高7.2厘米,口径15.6厘米。新疆维吾尔自治区察布查尔锡伯自治县出土。新疆维吾尔自治区文物考古研究所藏。此钵质地为夹砂红陶,敛口,斜腹,圆底,造型灵巧。内壁以红彩绘十字纹,外壁上部以黄彩绘树木纹和水波纹,是新疆地区出土陶器中的一件精致之作。

彩陶钵
新石器时代陶器。现藏辽宁博物馆。高8.5、腹径15、口径14.3、底径6厘米。手制,黄色细泥陶。大口、大腹、小平底。腹部有两小耳,口沿内圈及腹部绘有黑色斜三角形花纹带,下部无纹饰。