线粒体xianliti
广泛存在于真核细胞的细胞质中的一种由双层单位膜围成的细胞器,是细胞呼吸产生ATP的主要场所。最早发现线粒体的是R.A.科里凯尔(Kolliker)(1857),C.贝达(Benda)于1897年命名为线粒体。线粒体用詹那绿稀溶液活体染色后,在光学显微镜下即可看到。线粒体一般呈圆形、近圆形、棒状或线状,大小约0.3~0.8微米×0.4~3微米。细胞内线粒体的数目和分布与供能活动有关,消耗能量较多的细胞内线粒体数目多,细胞内需能部位线粒体比较集中。植物细胞内线粒体数目比动物细胞少,因线粒体的某些功能已被叶绿体取代。电镜下观察,线粒体由两层单位膜围成。外膜厚约6毫微米,蛋白质与脂类含量比约为1:1,膜的通透性很高。内膜厚约6~8毫微米,蛋白质与脂类含量比约为4:1,膜的通透性很低。内膜向内折叠成嵴,内膜和嵴的内表面上有许多有柄基粒。外膜上含有NADH—细胞色素C—还原酶系统,而内膜含有呼吸链和氧化磷酸化酶系。内外膜之间有宽约8.5毫微米的膜间腔,与嵴内腔构成一个连续空间,其中充满液体,含有腺苷酸激酶和核苷二磷酸激酶。内膜包围的线粒体内腔中充满基质,内有小的核糖体、磷酸钙沉淀颗粒,少量的环状DNA和RNA,以及三羧酸循环和脂肪酸氧化酶系等。线粒体是细胞呼吸的主要场所,三羧酸循环在线粒体基质中完成,通过呼吸链的氧化磷酸化在内膜上完成。在线粒体中葡萄糖、脂肪和氨基酸氧化分解释放的能量,以高能磷酸键形式贮存在ATP中,细胞生命活动需要的能量大约有95%来自线粒体中形成的ATP,故线粒体被称为化能转换器和细胞内能量供应的“动力工厂”。此外,线粒体还有贮存Ca24的作用。线粒体DNA能够复制,并有相应的蛋白质合成系统,但线粒体蛋白质大部分是由核控制。线粒体的增殖和生长是核基因和线粒体基因相互作用的结果。线粒体以分裂或出芽方式进行增殖。线粒体基因与细胞质遗传有一定关系。活细胞线粒体通过运动改变在细胞中的位置,不仅能够随着原生质流动而运动,而且能够向细胞需能较多的部位移动。线粒体基因指导合成蛋白质所用的遗传密码与核基因所用的密码有所不同。
线粒体mitochondria
真核细胞中进行氧化磷酸化作用并释放能量的半自主性的细胞器。
发现史 阿尔特曼(R.Altmann)于1894年首次发现细胞内有一种类似细菌大小的颗粒,称之为“原生粒”。1897年本达(C. Benda)将此种颗粒命名为线粒体。1913年沃伯格(O. Warburg)发现呼吸酶和线粒体之间有内在联系。1934年本斯利(R. Besley) 等从肝脏中分离出线粒体。1948年霍格布姆(G.H.Hogeboom)等确证线粒体是细胞的呼吸场所。60年代对线粒体的超微结构有了较深入的了解。70年代因发现线粒体是动物细胞内唯一含有DNA的细胞器,而被看作是细胞中的第二遗传系统。
形态 活体状态下,经詹纳斯绿B染色或用相差显微镜均可观察到线粒体。一般呈颗粒状或线状,也会出现棒状、哑铃状、球拍状和分支状等。直径0.5~1.0μm,长度一般为1~2μm,最长可达7μm。数量随细胞的大小和能量的需求而有很大差别,如精子细胞有20~24个,肝细胞1 000~1 600个,一种巨大多核变形虫可多达5×105个。绿色植物细胞线粒体比动物细胞少,因其部分能量由叶绿体提供。
分布 均匀分布于细胞质内,但也与需能状况有关,如在进行有丝分裂的细胞中,线粒体分布于纺锤丝之间,肌细胞的线粒体沿着肌原纤维排列,在肾小管细胞则集中于细胞基部靠近微血管处,植物细胞的线粒体可随原生质环流而移位。
结构 线粒体具内、外两层单位膜,厚为5~7nm。内外膜间有宽为6~8nm的膜间腔。外膜平整。内膜内包基质,常向基质内伸展形成片状或管状突起,称为嵴; 其内腔与膜内腔相通,称嵴内腔(图1、2)。嵴的形状和数量随物种、细胞类型和生活状态而变化,如肌细胞线粒体的嵴十分密集,而肝细胞和生殖细胞的嵴则较少。冬眠动物的细胞,不仅线粒体的数量增多,体积变大,嵴的密度也增高。通过低渗和负染可在嵴的内侧表面见到许多球状小颗粒,称基粒,其直径为8~9nm,间距约10nm,为合成腺苷三磷酸(ATP)的活性部位。每个线粒体有104~105个基粒。
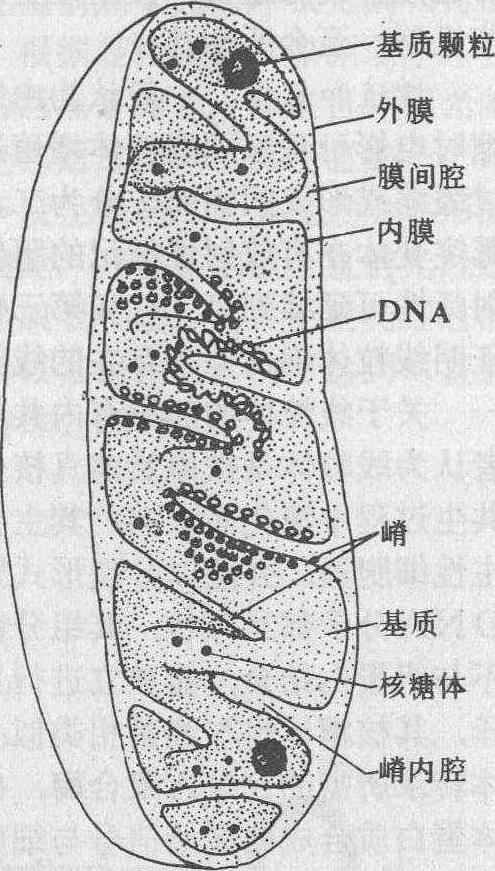
图 1 线粒体亚显微结构模式图
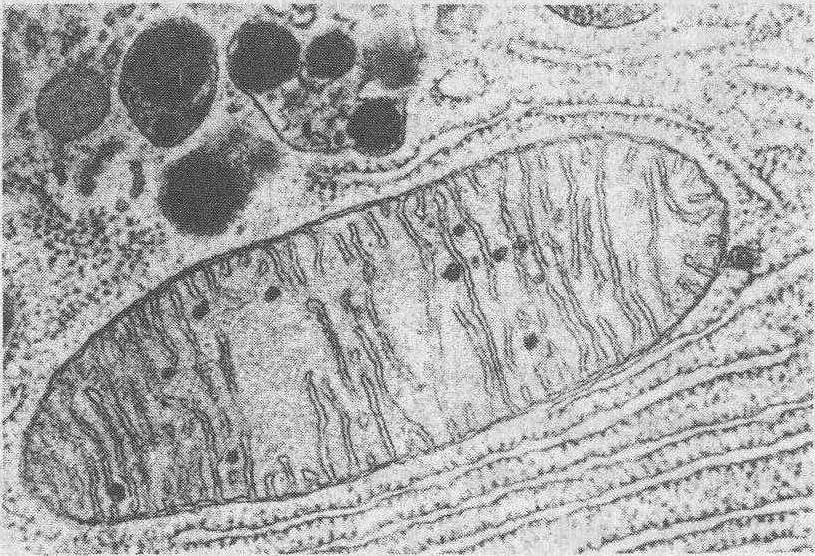
图 2 线粒体电镜图像
化学组成与功能
细胞的呼吸过程,除了糖酵解在细胞质中进行外,其余重要步骤如三羧酸循环,生物氧化和氧化磷酸化作用都在线粒体内完成,其中三羧酸循环的作用部位在基质,生物氧化和氧化磷酸化作用则在内膜上。
外膜 组成与细胞膜相似,脂类含量相对较高(蛋白质和脂类为1:1),磷脂主要为卵磷脂,其次为磷脂酰乙醇胺,磷脂酰肌醇较少,胆固醇量极微。通透性较大,水、电解质、蔗糖、尼克酰胺-磷酸核苷(NAD)和腺苷多磷酸(ATP和ADP)等及分子量小于1×104的质点都能自由通过。膜上还存在电压依赖阴离子选择通道,它是分子量3×104~3.5×104的蛋白质组成的孔道,表观直径2~4nm。阴离子(如Cl-)的通过速度较阳离子 (如K+、Na+) 为高,且当外膜电位较低(<20mV)时透性较大,较高时透性较小。其生物学意义在于呼吸作用的底物和辅助因子都是带负电荷或中性的物质,较大的有机阳离子如多胺化合物则为呼吸抑制剂,故外膜的这种选择透性有利于呼吸过程的正常进行,并且能通过内膜的生物氧化还原电位影响外膜的膜电位,从而调节呼吸作用的强度。
膜间腔 宽为6~8nm,与嵴内腔相通,腔内液体富含可溶性酶、底物和辅助因子。
内膜 蛋白质含量高(蛋白质和脂类为3.6:1),磷脂成分特殊,心磷脂含量超过20%,且有不少酸性磷脂; 不含胆固醇,但含有多量的醌类分子,约有50%的内膜蛋白是参与生物氧化和氧化磷酸化作用的酶和辅助因子,其余的膜蛋白则为各种物质的运输载体。内膜(以及外膜)上还有能识别由细胞质核糖体合成的线粒体组成蛋白质的特异受体,它能与该蛋白质前端的导肽结合,并根据导肽的信号而将该蛋白质分别分布在外膜、膜间腔、内膜内外侧或基质。
由于内膜的通透性比外膜小,很多物质均需依靠,特定的载体和通过不同的运输机制才能出入膜内基质。其中有些载体适应某种穿梭机制而起作用,如肉毒碱载体用于携带长链脂肪酸进入基质,而α-酮戊二酸和谷氨酸-天冬氨酸载体则用于将还原型尼克酰胺-腺嘌呤核苷(NADH)的还原力输入基质内,才能转化成ATP的高能键。
参与呼吸衔接作用的内膜蛋白自然形成5个酶复合体(见表),它们相互衔接、协调而组成生物氧化作用的电子传递链和氧化磷酸化作用的偶联因子。
线粒体内膜蛋白组成的复合体
| 名称 | 大 小 (×103) | 组成多肽链数目 | 每个小鼠肝细胞线 粒体中存在的数量 |
| Ⅰ Ⅱ Ⅲ Ⅳ Ⅴ | 700~900 140 250 160~170 500 | 25 4~5 9~10 8 12~14 | 350 700 1 000 2 000 1 000~1 600 |
复合体Ⅰ能使NADH氧化而还原泛醌(VQ);复合体Ⅱ能使琥珀酸氧化成延胡索酸而还原泛醌; 还原型泛醌能在内膜的脂相内游泳,而将质子(H+) 和电子(e-)传递给复合体Ⅲ,后者能使还原型泛醌氧化而使细胞色素C还原; 还原型细胞色素C在膜间腔内游泳,而将电子传递给复合体Ⅳ,后者能使还原型细胞色素C氧化而将氧分子还原成水。
由于复合体Ⅰ、Ⅲ、Ⅳ均能利用生物氧化作用产生的电能而作功,将基质内的质子(H+)主动运输到内膜外,起了质子泵的作用,其结果使内膜内侧质子浓度低于内膜外侧,质子便有从外向内扩散的被动运输趋势,即形成了质子动力,它具有作功的可能。当质子顺着浓度梯度由线粒体外经内膜上的复合体Ⅴ进入基质时便提供能量,使腺苷二磷酸(ADP)与无机磷酸合成ATP。因此,复合体V即为ATP合成酶系统。
复合体V亦称ATP酶复合体,分成基片(Fo)、柄(OSCP)和球头(F1)三部分。其中基片为内膜上的质子通道; 柄部为控制质子流的阀门; 球头则为利用质子动力合成ATP的活性部位,球头即电镜下所见的基本颗粒。复合体V亦是所谓偶联因子的生物学实体。
基质 在内膜围成的腔内含有许多可溶性蛋白,包括催化三羧酸循环、脂肪酸氧化和氨基酸降解等有关酶系,还有DNA、RNA、核糖体以及与DNA复制、转录有关的酶系。基质内含有少量致密颗粒,称基质颗粒,直径为20~60nm,是一些积累在线粒体内的钙离子和其他二价阳离子的磷酸盐颗粒。
植物细胞的线粒体与动物细胞的基本相同,但有不被氰化物或一氧化碳所抑制的电子传递系统,也不存在细胞色素C,却含有细胞色素b7。植物细胞的脂肪酸β氧化作用不在线粒体,而在乙醛酸循环体中进行。
线粒体的半自主性 线粒体具有完整的遗传系统,能进行遗传信息的复制、转录和翻译,但由于遗传信息量不够,大部分蛋白质仍需细胞核遗传系统提供,故线粒体只具有半自主性。多数动植物细胞的线粒体DNA都是裸露双螺旋环状分子,不与组蛋白结合,类似细菌DNA。一个线粒体中可含一至数个DNA分子,因此每个细胞中的线粒体DNA的数目很多,这些DNA分子的核苷酸顺序均相同。线粒体的遗传密码比较特殊,如UGA在线粒体中不是终止密码,而是蛋氨酸的密码和起始密码,AGA和AGG不代表精氨酸而是作为终止密码使用。玉米的细胞色素C氧化酶亚基Ⅱ的编码基因中UGG,CGG和UGA一样作为色氨酸的密码使用,故线粒体的遗传密码因生物种类的不同而有所差异。植物细胞线粒体的DNA含有植物雄性不育的基因,在其控制下,可合成控制雄蕊正常发育或雄性不育的一些蛋白质,农业上可用以测知植物不育系类型。
线粒体具有合成蛋白质的全套机构,包括mRNA、tRNA和核糖体。其分子量、沉降系数和核苷酸顺序均与核遗传系统不同,线粒体核糖体比较小,与细菌的相似,在基质内与内膜结合。对药物的敏感性也与核遗传系统不同,如氯霉素可抑制线粒体和细菌的蛋白质合成,而不抑制细胞质蛋白质合成; 环己亚胺能抑制细胞质蛋白质合成,但对线粒体和细菌却不起作用。线粒体中合成的蛋白质仅占线粒体蛋白质的10%,约10种左右,且多数是疏水性蛋白质。
由于线粒体DNA复制和基因表达所需的DNA聚合酶、核糖体蛋白质和各种氨基酸活化酶等均由核基因编码,在胞质核糖体上合成后再转移到线粒体,可能核-细胞质蛋白质合成系统就是通过合成这些物质来调节线粒体蛋白质合成系统的活性。另一方面线粒体也能合成阻遏物,控制核基因的表达。两者间的调节机制,至今所知甚少。
增殖和起源 一般认为由线粒体分裂而来。利用缩时电影可观察到线粒体增殖过程,它首先延长,通过缢缩或形成间隔而一分为二。将胆碱缺陷型的链孢霉突变体在有放射性标记的胆碱培养基中培养,其放射活性可延续到第二代和第三代,利用此种技术也可证明线粒体确由原来存在的线粒体产生。
关于线粒体的起源有内共生假说和分化假说。前者认为线粒体是被原始前真核生物吞噬的细菌在长期共生过程中演化而来的,其主要根据是线粒体是半自主性细胞器,并能以分裂形式繁殖; 线粒体与细菌的DNA分子都呈环状,其组分都缺5-甲基胞嘧啶,也不与组蛋白结合,能独立进行DNA复制、转录和翻译,其核糖体也与细菌相类似; 利福霉素能抑制线粒体转录所需的RNA聚合酶,也与细菌相类似; 线粒体蛋白质合成的起始信号与细菌相似,均是N-甲酰蛋氨酸,而不是真核细胞的蛋氨酸; 线粒体的内膜与原核细胞质膜相似,而外膜却与真核细胞相近,这些证据都支持了内共生发源说。分化假说认为线粒体是细胞进化过程中由质膜内陷而成。
线粒体
细胞(除细菌和蓝藻)中一种自体复制的细胞器。氧化磷酸化作用和ATP形成的场所。由一双层的膜所包被,膜的内层向内突起,把单个线粒体分隔成许多小室。细胞内线粒体总称线粒体系。
线粒体
是细胞质中的细胞器。在电子显微镜下,线粒体呈棒状或椭圆形小体,直径约0. 5微米,长约2~4微米。线粒体具有复杂的结构,外面由双层膜包绕,外层膜平滑,内层膜向内皱褶而形成许多线粒体嵴,嵴与嵴之间被分成许多相互连通的内室。在嵴的内表面覆盖着大量的球型颗粒,即酶系统,其中包括三羧循环酶系、脂肪酸氧化酶系、氧化磷酸化酶系。在适宜的条件下,它们能进行氧化磷酸化,以形成三磷酸腺苷(ATP)。因此,线粒体被称为细胞的能量“供应站”。线粒体担负着细胞呼吸的重要功能,它的完整,特别是线粒体嵴的存在,乃是氧化磷酸化不可缺少的条件。
线粒体mitochondrion
真核细胞内一种半自主的细胞器。包括3部分:
❶表面被外膜包围;
❷内有内膜,并向内褶叠伸出许多嵴,形成复杂的内部系统,嵴的内膜表面有ATP酶复合体;
❸内部系统的周围空间充满了基质。其功能主要是参与生物氧化、合成ATP,为细胞提供能量,供应细胞各项活动的需要,有“细胞动力站”之称。有自己的DNA、RNA、DNA聚合酶、RNA聚合酶、转移RNA、核糖体、氨基酸活化酶等所有合成蛋白质的一整套装置。在遗传上具有独立性、自主性,并在细胞中可自我复制和再生。
线粒体
真核细胞中细胞质内的粒状或棒状的细胞器。1894年由阿尔特曼(R.Altmann)首次发现。20世纪70年代发现线粒体是动物细胞内唯一含有DNA的细胞器。直径0.5~1.0 μm,长度1~2 μm,最长可达7 μm。具有内、外2层单位膜,厚为5~7 nm。内外膜间有宽为6~8 nm的膜间腔。外膜包围于外,内膜向内折叠成嵴,嵴膜内表面有基质颗粒,直径8~9 nm,间距约10 nm。每个线粒体约有104~105个基质颗粒。